第105页
- 第1页
- 第2页
- 第3页
- 第4页
- 第5页
- 第6页
- 第7页
- 第8页
- 第9页
- 第10页
- 第11页
- 第12页
- 第13页
- 第14页
- 第15页
- 第16页
- 第17页
- 第18页
- 第19页
- 第20页
- 第21页
- 第22页
- 第23页
- 第24页
- 第25页
- 第26页
- 第27页
- 第28页
- 第29页
- 第30页
- 第31页
- 第32页
- 第33页
- 第34页
- 第35页
- 第36页
- 第37页
- 第38页
- 第39页
- 第40页
- 第41页
- 第42页
- 第43页
- 第44页
- 第45页
- 第46页
- 第47页
- 第48页
- 第49页
- 第50页
- 第51页
- 第52页
- 第53页
- 第54页
- 第55页
- 第56页
- 第57页
- 第58页
- 第59页
- 第60页
- 第61页
- 第62页
- 第63页
- 第64页
- 第65页
- 第66页
- 第67页
- 第68页
- 第69页
- 第70页
- 第71页
- 第72页
- 第73页
- 第74页
- 第75页
- 第76页
- 第77页
- 第78页
- 第79页
- 第80页
- 第81页
- 第82页
- 第83页
- 第84页
- 第85页
- 第86页
- 第87页
- 第88页
- 第89页
- 第90页
- 第91页
- 第92页
- 第93页
- 第94页
- 第95页
- 第96页
- 第97页
- 第98页
- 第99页
- 第100页
- 第101页
- 第102页
- 第103页
- 第104页
- 第105页
- 第106页
- 第107页
- 第108页
- 第109页
- 第110页
- 第111页
- 第112页
- 第113页
- 第114页
- 第115页
- 第116页
- 第117页
- 第118页
- 第119页
- 第120页
- 第121页
- 第122页
- 第123页
- 第124页
- 第125页
- 第126页
- 第127页
- 第128页
- 第129页
- 第130页
- 第131页
- 第132页
- 第133页
- 第134页
- 第135页
- 第136页
- 第137页
- 第138页
- 第139页
- 第140页
- 第141页
- 第142页
- 第143页
- 第144页
- 第145页
- 第146页
- 第147页
- 第148页
- 第149页
- 第150页
- 第151页
- 第152页
- 第153页
- 第154页
- 第155页
- 第156页
- 第157页
- 第158页
- 第159页
- 第160页
- 第161页
- 第162页
- 第163页
- 第164页
- 第165页
- 第166页
- 第167页
- 第168页
- 第169页
- 第170页
- 第171页
- 第172页
- 第173页
- 第174页
- 第175页
- 第176页
- 第177页
- 第178页
- 第179页
- 第180页
- 第181页
- 第182页
- 第183页
- 第184页
- 第185页
- 第186页
- 第187页
- 第188页
- 第189页
- 第190页
- 第191页
- 第192页
- 第193页
- 第194页
- 第195页
- 第196页
- 第197页
- 第198页
- 第199页
- 第200页
- 第201页
- 第202页
- 第203页
- 第204页
- 第205页
- 第206页
- 第207页
- 第208页
- 第209页
- 第210页
- 第211页
- 第212页
- 第213页
- 第214页
- 第215页
- 第216页
- 第217页
- 第218页
- 第219页
- 第220页
- 第221页
- 第222页
- 第223页
- 第224页
- 第225页
- 第226页
- 第227页
- 第228页
- 第229页
- 第230页
- 第231页
- 第232页
- 第233页
- 第234页
- 第235页
- 第236页
- 第237页
- 第238页
- 第239页
- 第240页
- 第241页
- 第242页
- 第243页
- 第244页
- 第245页
- 第246页
- 第247页
- 第248页
- 第249页
- 第250页
- 第251页
- 第252页
- 第253页
- 第254页
- 第255页
- 第256页
- 第257页
- 第258页
- 第259页
- 第260页
- 第261页
- 第262页
- 第263页
- 第264页
- 第265页
- 第266页
- 第267页
- 第268页
- 第269页
- 第270页
- 第271页
- 第272页
- 第273页
- 第274页
- 第275页
- 第276页
- 第277页
- 第278页
- 第279页
- 第280页
- 第281页
- 第282页
- 第283页
- 第284页
- 第285页
- 第286页
- 第287页
- 第288页
- 第289页
- 第290页
- 第291页
- 第292页
- 第293页
- 第294页
- 第295页
- 第296页
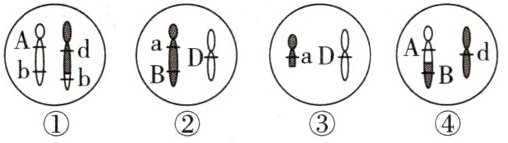
[例 3] 人类遗传病分为单基因遗传病、多基因遗传病和染色体异常遗传病,其中有很多类型的患儿都是父母在产生生殖细胞的过程中细胞异常所致。如图为某个体产生的四个精细胞,有关这些细胞的形成,下列分析正确的是(
[img]
A.①配子出现的原因是同源染色体中的非姐妹染色单体互换
B.①②③④四个精细胞可能由同一个精原细胞产生
C.④配子出现的原因是非同源染色体片段缺失所致
D.A、a 与 B、b 为非等位基因,在产生配子过程中遵循自由组合定律
B
)[img]
A.①配子出现的原因是同源染色体中的非姐妹染色单体互换
B.①②③④四个精细胞可能由同一个精原细胞产生
C.④配子出现的原因是非同源染色体片段缺失所致
D.A、a 与 B、b 为非等位基因,在产生配子过程中遵循自由组合定律
答案:
[例3] B [①配子上b基因所在片段与d基因所在片段结合,说明发生了非同源染色体的片段易位,而不是同源染色体中非姐妹染色单体的片段互换,A错误;从图中基因及染色体的关系来看,A、b基因在同一条染色体上,a、B基因在另一条与之同源的染色体上,而D、d基因位于另一对同源染色体上,①②③④可能来自同一个精原细胞(如图) ,B正确;④配子中,A基因所在片段与B基因所在片段结合,说明发生了同源染色体中非姐妹染色单体的片段互换,而不是非同源染色体的片段缺失,C错误;A、b基因在同一条染色体上,a、B基因在另一条与之同源的染色体上,所以A、a与B、b为非等位基因,在产生配子过程中不遵循自由组合定律,D错误。]
,B正确;④配子中,A基因所在片段与B基因所在片段结合,说明发生了同源染色体中非姐妹染色单体的片段互换,而不是非同源染色体的片段缺失,C错误;A、b基因在同一条染色体上,a、B基因在另一条与之同源的染色体上,所以A、a与B、b为非等位基因,在产生配子过程中不遵循自由组合定律,D错误。]
[例3] B [①配子上b基因所在片段与d基因所在片段结合,说明发生了非同源染色体的片段易位,而不是同源染色体中非姐妹染色单体的片段互换,A错误;从图中基因及染色体的关系来看,A、b基因在同一条染色体上,a、B基因在另一条与之同源的染色体上,而D、d基因位于另一对同源染色体上,①②③④可能来自同一个精原细胞(如图)
,B正确;④配子中,A基因所在片段与B基因所在片段结合,说明发生了同源染色体中非姐妹染色单体的片段互换,而不是非同源染色体的片段缺失,C错误;A、b基因在同一条染色体上,a、B基因在另一条与之同源的染色体上,所以A、a与B、b为非等位基因,在产生配子过程中不遵循自由组合定律,D错误。] [例 4] (不定项)为探究一定浓度的西洛酰胺(Cil)和$0.5ng/mL$的雌二醇(E2)对绵羊卵原细胞减数分裂不同时期的比例及环腺苷酸(cAMP)浓度的影响,某科研小组进行了相关实验,一段时间后,测得实验结果如表。下列有关叙述或推测,错误的是(
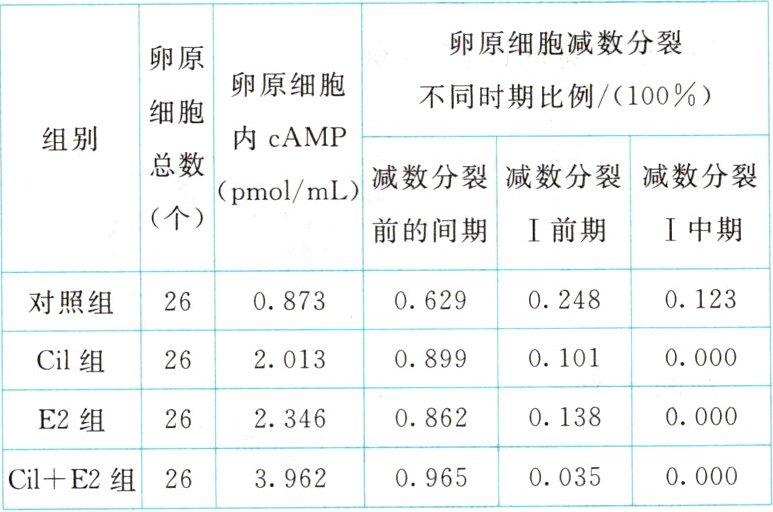
A.减数分裂前的间期,染色体复制后,染色体数目加倍
B.若降低细胞内 cAMP 浓度,同源染色体不能进行联会
C.一定浓度的 Cil 可能促进卵原细胞内 cAMP 的合成
D.$0.5ng/mL$的 E2 可阻滞减数分裂进程
AB
)A.减数分裂前的间期,染色体复制后,染色体数目加倍
B.若降低细胞内 cAMP 浓度,同源染色体不能进行联会
C.一定浓度的 Cil 可能促进卵原细胞内 cAMP 的合成
D.$0.5ng/mL$的 E2 可阻滞减数分裂进程
答案:
[例4] AB [减数分裂前的间期,染色体复制后,核DNA加倍,染色体数目没有加倍,A错误;与对照组相比,Cil组和E2组的cAMP浓度较高,处于减数分裂前的间期的细胞数较多,推测Cil和E2可通过提高cAMP浓度阻滞卵原细胞减数分裂,故降低cAMP浓度,进入减数分裂的卵原细胞较多,同源染色体能进行联会,B错误;与对照组相比,Cil组的卵原细胞内cAMP浓度较高,故一定浓度的Cil可能促进卵原细胞内cAMP的合成,C正确;与对照组相比,E2组中处于减数分裂I中期的细胞比例为0,故0.5ng/mL的E2可阻滞减数分裂进程,D正确。]
[例 5] 某种昆虫的性别由 X 染色体的数量决定,雌性个体的体细胞中有 12 条染色体,性染色体为两条 X 染色体;雄性个体的体细胞中有 11 条染色体,性染色体为一条 X 染色体,在减数分裂Ⅰ后期,复制后的 X 染色体随机移向细胞一极。在自然状态下,该种昆虫种群的性别比例为$1:1$。不考虑变异,下列说法正确的是(
A.该昆虫的初级精母细胞在减数分裂Ⅰ前期形成 6 个四分体
B.若仅考虑染色体组成,该昆虫的雄性个体产生的精子种类少于$2^5$种
C.该昆虫的一个次级精母细胞中,减数分裂Ⅱ后期时移向两极的染色体数量不相等
D.该种昆虫种群的性别比例为$1:1$的原因之一是含 X 染色体的精子与不含 X 染色体的精子比例相等
D
)A.该昆虫的初级精母细胞在减数分裂Ⅰ前期形成 6 个四分体
B.若仅考虑染色体组成,该昆虫的雄性个体产生的精子种类少于$2^5$种
C.该昆虫的一个次级精母细胞中,减数分裂Ⅱ后期时移向两极的染色体数量不相等
D.该种昆虫种群的性别比例为$1:1$的原因之一是含 X 染色体的精子与不含 X 染色体的精子比例相等
答案:
[例5] D [四分体是复制后的同源染色体相互配对后形成的,分析题意可知,该类昆虫雄性个体的体细胞中有11条染色体,5对同源染色体+1条X染色体,故减数分裂I前期形成5个四分体,A错误;该昆虫的雄性个体有11条染色体,若仅考虑5对同源染色体,则该昆虫的雄性个体产生的精子种类有$2^{5}=32$种,且在减数分裂I后期,复制后的X染色体随机移向细胞一极,故该昆虫的雄性个体产生的精子种类大于32种,B错误;若不考虑染色体变异,该昆虫的一个次级精母细胞中,由于着丝粒的分裂,减数分裂II后期时移向两极的染色体数量相等,C错误;由于在减数分裂I后期,复制后的X染色体随机移向细胞一极,故该种昆虫种群的性别比例为1:1的原因之一是含X染色体的精子与不含X染色体的精子比例相等,D正确。]
查看更多完整答案,请扫码查看

